Передвижение воды по растению. Пути и механизмы передвижения воды по растению
Основным источником влаги является вода, находящаяся в почве, и основным органом поглощения воды в растение - корневая система. Роль этого органа, прежде всего, заключается в том, что благодаря огромной поверхности обеспечивается поступление воды в растение из большего объема почвы.
Сформировавшаяся корневая система представляет собой сложный орган с хорошо дифференцированной внешней и внутренней структурой. Корневая система имеет поглощающую или всасывающую зону - это зона корневых волосков. Поступив в клетку корневого волоска, вода становится частью живой системы - клетки растения - и подчиняется закономерностям, действующим в живой клетке.
Передвижение по растению определяется двумя основными двигателями водного потока в растении: нижним двигателем водного потока или корневым давлением, верхним двигателем водного потока или присасывающим действием атмосферы.
Основной силой, вызывающей поступление и передвижение воды в растении, является процесс транспирации, в результате которого возникает градиент водного потенциала. Градиент водного потенциала между клеткой и окружающим пространством создает движущую силу потока воды через мембрану. Если окружающая клетку среда представляет собой гипертонические, более концентрированные, чем клеточный сок, растворы, то вода станет выходить из клетки наружу. Это приведет к потере тургора клеткой, отделению плазмалеммы от клеточной стенки и обособлению протопласта - явлению плазмолиза.
Механизм, обеспечивающий поднятие воды по растению за счет корневого давления, - носит название нижнего концевого двигателя водного тока.
Корневое давление создается при переходе воды из коры корня в сосудистую систему корня при прохождении воды через пропускные клетки перицикла, из которых вода под давлением как бы впрыскивается в сосуды ксилемы. Доказательством этого служат явления гуттации и «плача растений».
Вода, поглощенная корневыми волосками и другими клетками эпидермиса, из клеток внешней части корня перемещается к ксилеме, занимающей центральную часть корня. Главным путем диффузии воды во внешней части корня служит аполаст - непрерывная совокупность клеточных стенок. Однако в эндодерме (цилиндрическом слое клеток, окружающем проводящую ткань) свободная диффузия по клеточным стенкам наталкивается на преграду - водонепроницаемый пробковый слой пояска Каспари. Вода должна изменить здесь свой путь и пройти сквозь мембрану и протопласт клеток эндодермы, играющей, таким образом, роль осмотического барьера между корой корня и его центральным цилиндром. У однодольных пробковеют также и внутренние тангенциальные стенки клеток, но эти стенки пронизаны порами, по которым, как по каналам, может проходить вода.
По ксилеме вода поднимается в надземные части растения. Ксилема состоит из нескольких типов клеток. Вода движется в ней главным образом по сосудам и трахеидам. И те и другие клетки прекрасно приспособлены для этой цели: они вытянуты в длину, лишены живого содержимого и внутри полые, т. е. это как бы трубки для воды. Одревесневшие вторичные клеточные стенки достаточно прочны на разрыв, чтобы выдерживать огромную разность давлений, возникающую при подъеме воды к вершинам высоких деревьев. Торцевые, а иногда и боковые стенки члеников сосудов перфорированы; сосуды, состоящие из соединенных конец в конец члеников, образуют длинные трубки, по которым легко проходит вода с растворенными в ней минеральными веществами. В трахеидах нет перфораций, и вода, для того чтобы попасть из одной тра-хеиды в другую, должна пройти через их торцевые стенки; однако трахеиды - очень длинные клетки, а потому и эта конструкция достаточно хорошо приспособлена для проведения воды.
У цветковых растений есть и сосуды, и трахеиды; у более примитивных форм сосудов, как правило, нет.
Листовые жилки, состоящие из тяжей ксилемы и флоэмы, образуют в листе настолько густую сеть, что любая его клетка оказывается достаточно близко от источника воды. Из ксилемы вода диффундирует в стенки клеток мезофилла. Таким образом, вода в жидкой фазе заполняет весь путь от почвы - через корень и стебель - до клеток мезофилла в листе. Суммарный поток воды направлен всегда в сторону меньшего водного потенциала, т. е. максимален в почве, несколько ниже в клетках корня и самый низкий в клетках, примыкающих к эпидермису листа. Малая величина показателя в этих последних клетках объясняется главным образом испарением воды с поверхности листа.
Вода переходит из растения в окружающий воздух главным образом в парообразном состоянии. В мезофилле листа имеются обширные межклеточные пространства, и каждая клетка мезофилла хотя бы одной своей стороной граничит с таким межклетником. Вследствие испарения воды с влажных клеточных стенок воздух в межклетниках насыщен водяными парами, и часть этих паров теряется - выходит наружу. Поскольку у большинства растений клетки эпидермиса покрыты воскообразной водонепроницаемой кутикулой, водяные пары выходят из листа в атмосферу главным образом через устьица.
Гуттация - это выделение капельно-жидкой влаги листьями через гидатоды в условиях затрудненного испарения. Плач растения - это вытекание пасоки (воды с растворенными в ней минеральными веществами, находящейся в ксилеме) из стеблей растений со срезанными побегами. Гуттация обычно имеет место в условиях высокой влажности воздуха, когда «выключен»верхний концевой двигатель водного токатранспирация - физиологический процесс испарения воды надземными органами растений. Он осуществляется в основном из листьев через устьица и кутикулу.
Сначала, прочитав учебник, энциклопедии и статьи в Интернете, я узнала, как выглядит строение цветочных растений
Органы цветочного растения - побег, корень, цветки, плоды с семенами. И все они состоят из различных видов ткани: образовательной, покровной, механической, проводящей, основной. Все эти ткани выполняют различные функции в жизни растений.
Транспорт воды в цветочных растениях
Чтобы растение могло расти, должны выполняться определенные условия: свет, тепло, вода, питание. Активное перемещение веществ у растений происходит по проводящим тканям. Вода и растворенные в ней минеральные вещества передвигаются в растении от корней к цветку по сосудам. Вода поступает в растение через корневые волоски, затем вода по сосудам корня под давлением поднимается. Попав в листья, вода испаряется с поверхности клеток и в виде пара выходит в атмосферу. Этот процесс обеспечивает непрерывный восходящий ток воды по растению.
Но какие силы обеспечивают движение тока воды вверх по стеблю в стакане с водой? Можно предположить, что вода выталкивается снизу или ее тянут сверху. На небольшие расстояния транспорт веществ обеспечивают физические процессы диффузии. Молекулы воды передвигаются из той области, где их концентрация высока, туда, где их концентрация низка.
1.3 Передвижение воды по растению
Вода, поглощенная клетками корня, под влиянием разности водных потенциалов, которые возникают благодаря транспирации, а также силе корневого давления, передвигается до проводящих путей ксилемы. Согласно современным представлениям, вода в корневой системе передвигается не только по живым клеткам. Еще в 1932 г. немецкий физиолог Мюнх развил представление о существовании в корневой системе двух относительно не зависимых друг от друга объемов, по которым передвигается вода, - апопласта и симпласта. Апопласт – это свободное пространство корня, в которое входят межклетные промежутки, оболочки клеток, а также сосуды ксилемы. Симпласт – это совокупность протопластов всех клеток, отграниченных полупроницаемой мембраной. Благодаря многочисленным плазмодесмам, соединяющим между собой протопласт отдельных клеток, симпласт представляет единую систему. Апопласт, по-видимому, не непрерывен, а разделен на два объема. Первая часть апопласта расположена в коре корня до клеток эндодермы, вторая – по другую сторону клеток эндодермы, и включает в себя сосуды ксилемы. Клетки эндодермы благодаря пояскам Каспари представляют как бы барьер для передвижения воды по свободному пространству (межклетникам и клеточным оболочкам). Для того чтоб попасть в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану и главным образом по апопласту и лишь частично по симпласту. Однако в клетках эндодермы передвижение воды идет, по-видимому, по симпласту. Далее вода поступает в сосуды ксилемы. Затем передвижение воды идет по сосудистой системе корня, стебля и листа.
Из сосудов стебля вода движется через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Иногда мелких ответвлений жилок листа так много, что они подводят воду почти к каждой клетке. Вся вода в клетке находится в равновесном состоянии. Иначе говоря, в смысле насыщенности водой, имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. Вода передвигается от клетки к клетке благодаря градиенту сосущей силы.
Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту значительно большую 10 м. сила сцепления увеличивается, так как молекулы воды обладают большим сродством друг к другу. Силы сцепления обладают и между водой и стенками сосудов.
Степень натяжения водных нитей в сосудах зависит от соотношения процессов поглощения и испарения воды. Все это позволяет растительному организму поддерживать единую водную систему и не обязательно восполнять каждую каплю испаряемой воды.
В том случае, если в отдельные членики сосудов попадает воздух, они, по-видимому, выключается из общего тока проведения воды. Таков путь передвижения воды по растению (рис. 2).
Рис.2. Путь воды в растении.
Скорость перемещения воды по растению в течение суток изменяется. В дневные часы она на много больше. При этом разные виды растений различаются по скорости передвижения воды. Изменение температуры, введение метаболических ингибиторов не влияют на передвижение воды. Вместе с тем этот процесс, как и следовало ожидать, очень сильно зависит от скорости транспирации и от диаметра водопроводящих сосудов. В более широких сосудах вода встречает меньшее сопротивление. Однако надо учитывать, что в более широкие сосуды могут попасть пузырьки воздуха или произойти какие-либо иные нарушения тока воды.
2.Транспирация: физиологические механизмы
В основе расходования воды растительным организмом лежит физиологический процесс испарения – переход воды из жидкого в парообразное состояние, происходящей при соприкосновении органов растения с не насыщенной водой атмосферой. Однако этот процесс осложнен физиологическими и анатомическими особенностями растения, и его называют транспирацией
2.1 Назначение транспирации
В обычно протекающих процессах транспирация не является необходимой. Так если выращивать растения в условиях высокой и низкой влажности воздуха, то, естественно, в первом случае транспирация будет идти сознательно меньшей интенсивностью. Однако рост растений будет одинаков или даже лучше там, где влажность воздуха выше, а транспирация меньше. Известно, что большая часть всей поглощенной энергии тратится на транспирацию, которая в определенном объеме полезна растительному организму.
1.Транспирация спасает растение от перегрева, который ему грозит на прямом солнечном свете. Температура сильно транспирирующего листа может быть примерно 7`C ниже температуры листа завязающего, не транспирирующего. Это особенно важно в связи с тем, что перегрев, разрушая хлоропласты, резко снижает процесс фотосинтеза (оптимальная температура для процесса фотосинтеза около 30-33`C). Именно благодаря высокой транспирирующей способности многие растения хорошо переносят повышенную температуру.
2.Транспирация создает непрерывный ток воды из корневой системы к листьям, который связывает все органы растения в единое целое.
3. С транспирационным током передвигаются растворимые минеральные и частично органические питательные вещества, при этом, чем интенсивнее транспирация, тем быстрее идет процесс передвижения.
2.2 Лист как орган транспирации
Основным транспортирующим органом является лист. Средняя толщина листа составляет 100-200 мкм. Паренхимных клетки листа расположены рыхло, между ними имеется система межклетников, составляющая в общей сложности от 15 до 25% объема листа. Лист окружен покровной тканью- эпидермисом, состоящим из компактно расположенных клеток, наружные стенки которых утолщены. Листья большинства растений покрыта кутикулой, в состав которой входит оксимонокарбоновые кислоты, содержащие по 16-18 атомов углерода и по 2-3 гидроксильных группы. Эти кислоты соединены друг с другом в цепочки с помощью эфирных связей. Кутикула варьирует как по составу, так и по толщине. Более развитой кутикулой характеризуются листья светолюбивых растений по сравнению с влаголюбивыми. Кутикула вместе с клетками эпидермиса образует как бы барьер на пути испарения паров воды. При этом особенно значительную преграду составляет кутикула. Удаление кутикулы во много раз повышает интенсивность испарения. Сопротивление выходу паров оказывают в определенной мере и утолщенные стенки клетки эпидермиса. Все эти особенности выработались в процессе эволюции как приспособление к сокращению испарения. Для соприкосновения листа с атмосферой имеются устьица. Устьица – одно из оригинальных приспособлений листа, обладающее способностью открываться и закрываться. Обычно устьичные отверстия ограничены двумя замыкающими клетками, стенки которых не равномерно утолщены. У двудольных растений замыкающие клетки бобовидной, или полулунной, формы, при этом их внутренние прилегающие друг к другу стенки долее толстые, а внешние – более тонкие. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на стенки и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель. У однодольных растений строение замыкающих клеток несколько иное. Они представлены двумя удлинёнными клетками, на концах которых стенки более тонкие. При насыщении водой более тонкие стенки на концах растягиваются и раздвигают замыкающие клетки, благодаря чему образуется щель. (рис.3)

Рис.3 Структура устьиц у двудольных (А) и однодольных (Б) растений:
1-устьичная щель; 2-ядро; 3-хлоропласты; 4-толстая клеточная стенка; 5-замыкающие клетки устьиц; 6-побочные клетки; 7-клетки эпидермиса с многочисленными порами.
Число устьичных отверстий колеблется в зависимости от вида растения от 1 до 60 тыс. на 1 кв.см. листа. Большая часть устьиц расположена на нижней стороне листа. Диаметр устьичных щелей составляет всего 3-12 мкм.
Устьица соединяют внутренние пространство листа с внешней средой. Вода проступает в лист через сеть жилок, в которых расположены сосудистые элементы. Возможны два пути испарения: 1) через наружные стенки клеток эпидермиса в атмосферу; 2) через стенки клеток мезофилла в межклеточное пространство листа и далее в парообразном состоянии через устьица. В связи с этим различают устьичную и кутикулярную транспирацию. В том, что действительно испарение идет не только через устьица, но и через кутикулу, легко убедиться. Так, если взять листья, у которых устьица расположены только с нижней стороны (например, листья яблони), и замазать эту сторону вазелином, то испарение воды будет продолжаться, хотя и в значительно уменьшенном размере. Следовательно, определенное количество воды испаряется через кутикулу.
Кутикулярная транспирация обычно составляет около 10% от общей потери воды листом. Однако в некоторых случаях у растений, листья которых характеризуются слабым развитием кутикулы, доля этого вида транспирации может повышаться до 30%. Имеет значение также возраст листа. Молодые листья, как правило, имеют слабо развитую кутикулу и, следовательно, более интенсивную кутикулярную транспирацию. Наименьшая Кутикулярная транспирация наблюдается у листьев, Закончивших свой рост. У старых листьев доля кутикулярной транспирации снова возрастает, так как, хотя кутикула и сохраняет достаточную толщину, в ней появляются трещины, через которые легко проходят пары воды.
Все же основная часть воды испаряется через устьица. Процесс устьичной транспирации можно подразделить на ряд этапов.
Первый этап – это переход воды из клеточных оболочек, где она находится в капельно-жидком состоянии, в межклетники (парообразное состояние). Это собственно процесс испарения. Важно подчеркнуть, что уже на этом этапе растение обладает способностью регулировать процесс транспирации (внеустьичная регулировка). Это связано с несколькими причинами: 1. Между всеми частями клетки существует водное равновесие. Чем меньше воды в клетке, тем выше становится концентрация клеточного сока. А это, в свою очередь, будет уменьшать интенсивность испарения. 2. Между микро- и макро- фибриллами целлюлозы, составляющими клеточные оболочки, имеются капиллярные промежутки. Вода испаряется именно из капилляров. Когда воды в клетках достаточно, клеточные оболочки насыщенны водой, мениски в капиллярах имеют выпуклую форму, силы поверхностного натяжения ослаблены. В этом случае молекулы воды легко отрываются и переходят в парообразное состояние, заполняя межклетники. При уменьшении содержания воды мениски в капиллярах становятся более вогнутыми, это увеличивает силы поверхностного натяжения, и вода с большей силой удерживается в клеточных оболочках. Чем более вогнут мениск, тем путь молекул воды до межклеточных пространств более длинен и извилист. В результате интенсивность испарения сокращается. Таким образом, уже на этом первом этапе растение испаряет тем меньше воды, чем меньше её содержит.
Второй этап – это выход паров воды из межклетников через устьичные щели. Поверхность всех клеточных стенок, соприкасающихся с межклетными пространствами, повышает поверхность листа примерно в 10-30 раз. Все же если устьица закрыты, то все это пространство быстро насыщается парами воды и переход воды из жидкого в парообразное состояние прекращается. Иная картина наблюдается при открытых устьицах. Как только часть паров воды выедет из межклетников через устьичные щели, так сейчас же этот недостаток восполняется за счет испарения воды с поверхности клеток. Поэтому степень открытости устьиц является основным механизмом, регулирующим интенсивность транспирации. При открытых устьицах общая поверхность устьичных щелей составляет всего 1-2% от площади листа. Казалось бы, это должно очень сильно уменьшать испарение по сравнению с испарением свободной водной поверхности той же площади, что и лист. Однако это не так. Сравнение испарения листа с испарением со свободной водной поверхности той же площади показало, что оно идет не в 100 раз, как это следовало бы, исходя из размеров открытой площади (1%), а всего в два раза медленнее. Объяснение этому явлению было дано в исследованиях английских физиологов Брауна и Эскомба, которые установили, что испарение из ряда мелких отверстий идет быстрее, чем из одного крупного той же площади. Это связано с так называемым явлением краевой диффузии. При диффузии из отверстий, отстоящих друг от друга на некотором расстоянии, молекулы воды, расположенные по краям, рассеиваются быстрее. Естественно, что таких краевых молекул значительно больше в ряде мелких отверстий по сравнению с одним крупным. В связи с этим для малых отверстий интенсивность испарения пропорциональна их диаметру, а не площади. Это видно из данных таблицы.
| Диаметр пор, | Площадь пор, В отн.ед. (пd 2) | Периметр, | Испарение воды, |
Указанная закономерность проявляется в том случае, если мелкие поры расположены достаточно далеко друг от друга. Структура листа удовлетворяет указанным требованиям. Поры (устьица) имеют малый диаметр и достаточно удалены друг от друга. При открытых устьицах выход паров воды идет достаточно интенсивно, закрытие устьиц резко тормозит испарение. Именно на этом этапе вступает в действие устьичная регулировка транспирации. При недостатке воды в листе устьица автоматически закрываются.
Полное закрытие устьиц сокращает транспирацию примерно на 90%. Вместе с тем уменьшение диаметра устьичных щелей не всегда приводит к соответственному сокращению транспирационного процесса. Определения показали, что устьица должны закрываться больше чем на Ѕ, для того чтобы это сказалось на уменьшении интенсивности транспирации.
Третий этап транспирации – это диффузия паров воды от поверхности листа в более далекие слои атмосферы. Этот этап регулируется лишь условиями внешней среды.
3.Адаптация к дефициту воды
Вода является необходимым условием существования всех живых организмов на Земле. Значение воды в процессах жизнедеятельности определяется тем, что она является основной средой в клетке, где осуществляются процессы метаболизма, служит важнейшим исходным, промежуточным или конечным продуктом биохимических реакций. Особая роль воды для наземных организмов (особенно растений) заключается в необходимости постоянного пополнения ее, из-за потерь при испарении. Поэтому вся эволюция наземных организмов шла в направлении приспособления к активному добыванию и экономному использованию влаги. Наконец, для многих видов растений, животных, грибов и микроорганизмов вода является непосредственной средой их обитания.
Увлажненность местообитания и, как следствие, водообеснечение наземных организмов зависят, прежде всего, от количества атмосферных осадков, их распределения по временам года, наличия водоемов, уровня грунтовых вод, запасов почвенной влаги и тд. Влажность оказывает влияние на распространение растений и животных, как в пределах ограниченной территории, так и в широком географическом масштабе, определяя их зональность (смена лесов степями, степей - полупустынями и пустынями).
При изучении экологической роли воды учитывается не только количество выпадающих осадков, но и соотношение их величины и испаряемости. Области, в которых испарение превышает годовую величину суммы осадков, называются аридными (сухими, засушливыми). В аридных областях растения испытывают недостаток влаги в течение большей части вегетационного периода. В гумидпых (влажных) областях растения обеспечены водой в достаточной мере.
Экологические группы растений по отношению к влаге и их адаптации к водному режиму. Высшие наземные растения, ведущие прикрепленный образ жизни, в большей степени, чем животные, зависят от обеспеченности субстрата и воздуха влагой. По приуроченности к местообитаниям с разными условиями увлажнения и по выработке соответствующих приспособлений среди наземных растений различают три основные экологические группы: гигрофиты, мезофиты и ксерофиты. Условия водоснабжения существенно влияют на их внешний облик и внутреннюю структуру.
Гигрофиты - растения избыточно увлажненных местообитаний с высокой влажностью воздуха и почвы. Для них характерно отсутствие приспособлений, ограничивающих расход воды, и неспособность переносить даже незначительную ее потерю. Наиболее типичные гигрофиты - травянистые растения и эпифиты влажных тропических лесов и нижних ярусов сырых лесов в разных климатических зонах (чистотел большой, недотрога обыкновенная, кислица обыкновенная и др.), прибрежные виды (калужница болотная, плакун-трава, рогоз, камыш, тростник), растения сырых и влажных лугов, болот (белокрыльник болотный, сабельник болотный, вахта трехлистная, осоки), некоторые культурные растения.
Характерные структурные черты гигрофитов - тонкие листовые пластинки с небольшим числом широко открытых устьиц, рыхлое сложение тканей листа с крупными межклетниками, слабое развитие водопроводящей системы (ксилемы), тонкие слаборазветвленные корни, часто без корневых волосков. К физиологическим адаптациям гигрофитов следует отнести низкое осмотическое давление клеточного сока, незначительную водоудерживающую способность и, как следствие, высокую интенсивность транспирации, которая мало отличается от физического испарения. Избыточная влага удаляется также путем гуттации - выделения воды через специальные выделительные клетки, расположенные по краю листа. Избыточная влага затрудняет аэрацию, а следовательно, дыхание и всасывающую деятельность корней, поэтому удаление излишков влаги представляет собой борьбу растений за доступ воздуха.
Ксерофиты - растения сухих местообитаний, способные переносить продолжительную засуху, оставаясь физиологически активными. Это растения пустынь, сухих степей, саванн, сухих субтропиков, песчаных дюн и сухих, сильно нагреваемых склонов. Структурные и физиологические особенности ксерофитов нацелены на преодоление постоянного или временного недостатка влаги в почве или воздухе. Решение данной проблемы осуществляется тремя способами:
1) эффективным добыванием (всасыванием) воды
2) экономным ее расходованием
3) способностью переносить большие потери воды
Интенсивное добывание воды из почвы достигается ксерофитами благодаря хорошо развитой корневой системе. По общей массе корневые системы ксерофитов примерно в 10 раз, а иногда и в 300-400 раз превышают надземные части. Длина корней может достигать 10-15 м, а у саксаула черного - 30-40 м, что позволяет растениям использовать влагу глубоких почвенных горизонтов, а в отдельных случаях и грунтовых вод. Встречаются и поверхностные, хорошо развитые корневые системы, приспособленные к поглощению скудных атмосферных осадков, орошающих лишь верхние горизонты почвы.
Экономное расходование влаги ксерофитами обеспечивается тем, что листья у них мелкие, узкие, жесткие, с толстой кутикулой, с многослойным толстостенным эпидермисом, с большим количеством механических тканей, поэтому даже при большой потере воды листья не теряют упругости и тургора. Клетки листа мелкие, плотно упакованы, благодаря чему сильно сокращается внутренняя испаряющая поверхность. Кроме того, у ксерофитов повышенное осмотическое давление клеточного сока, благодаря чему они могут всасывать воду даже при больших водоотнимающих силах почвы.
К физиологическим адаптациям относится и высокая водоудерживающая способность клеток и тканей, обусловленная большой вязкостью и эластичностью цитоплазмы, значительной долей связанной воды в общем водном запасе и т. д. Это позволяет ксерофитам переносить глубокое обезвоживание тканей (до 75% всего водного запаса) без потери жизнеспособности. Кроме того, одной из биохимических основ засухоустойчивости растений является сохранение активности ферментов при глубоком обезвоживании.
Ксерофиты с наиболее ярко выраженными ксероморфными чертами строения листьев, перечисленными выше, имеют своеобразный внешний облик, за что получили название склерофиты.
К группе ксерофитов относятся и суккуленты - растения с сочными мясистыми листьями или стеблями, содержащими сильно развитую водоносную ткань. Различают листовые суккуленты (агавы, алоэ, молодило, очитки) и стеблевые, у которых листья редуцированы, а надземные части представлены мясистыми стеблями (кактусы, некоторые молочаи и др.). Фотосинтез у стеблевых суккулентов осуществляется периферическим слоем паренхимы стебля, содержащим хлорофилл. Длительные засушливые периоды преодолеваются ими путем накопления воды в водоносных тканях, связывания ее коллоидами клеток, экономного расходования, которое обеспечивается защитой эпидермиса растений восковым налетом, погруженными в ткань листа или стебля немногочисленными днем закрытыми устьицами. В результате транспирация у суккулентов чрезвычайно мала: в пустынях кактусы из рода Camegia транспирируют в сутки всего лишь I -3 мг воды на 1 г сырой массы.
Корневая система поверхностная, мало развитая, рассчитана на поглощение воды из верхних слоев почвы, увлажненных редко выпадающими дождями. В засуху корни могут отмирать, но после дождей быстро (за 2-4 дня) отрастают новые. Суккуленты приурочены главным образом к засушливым зонам Центральной Америки, Южной Африки, Средиземноморья.
Мезофиты - занимают промежуточное положение между гигрофитами и ксерофитами. Они распространены в умеренно влажных зонах с умеренно теплым режимом и достаточно хорошей обеспеченностью минеральным питанием. К мезофитам относятся растения лугов, травянистого покрова лесов, лиственные деревья и кустарники из областей умеренно влажного климата, а также большинство культурных растений и сорняки. Для мезофитов характерна высокая экологическая пластичность, позволяющая им адаптироваться к меняющимся условиям внешней среды.
Специфичные пути регуляции водообмена позволили растениям занять самые различные по экологическим условиям участки суши. Многообразие способов приспособления лежит, таким образом, в основе распространения растений на Земле, где дефицит влаги является одной из главных проблем экологической адаптации.
Заключение
Из всего выше перечисленного можно вывести общее заключение, что при дефиците влаги растения могут адаптироваться е.т. образование корневой системы, которая достигает влажных зон почвы; ограничение расхода воды на транспирацию; запасание воды в тканях растений.
Так как вода является основной составной частью растительных организмов. Вода- это та среда, в которой протекает все процессы обмена веществ.
Водный ток обеспечивает связь между отдельными органами растений. Питательные вещества передвигаются по растению в растворенном виде. Насыщенность водой (тургор) обеспечивает прочность тканей, сохранение структуры травянистых растений, определенную ориентировку организмов растений в пространстве. Рост клеток в фазе растяжения идет главным образом за счет накопления воды в вакуоли.
Таким образом, вода обеспечивает протекание процессов обмена, коррелятивные взаимодействия, связь организма со средой. Для нормальной жизнедеятельности клетка должна быть насыщенна водой.
Основным источником влаги является вода, находящаяся в почве, и основным органном поглощения воды является корневая система. Роль этого органа прежде всего заключается в том, что благодаря огромной поверхности обеспечивается поступление воды в растения из возможно большого объема почвы. Сформировавшаяся корневая система представляет собой сложный орган с хорошо дифференцированной внешней и внутренней структурой.
Литература
1.Якушкина Н.И. Физиология растений. М., Просвещение, 1980г.
2.Козловский Водный обмен растений. М., Колос. 1969г.
3.Сказкин Ф.Д. Критический период у растений к недостаточному водоснабжению. М., Наука. 1968г.
4.Радкевич В.А. Экология. Мин., Высшая школа. 1983г.
5.Генкель П.А. Физиология устойчивости растительных организмов. М., Изд-во МГУ. 1967г. т.3.
6. Жданов В.С. Аквариумные растения. / Под. ред. Коровина. -
М.: Лесн. пром-ть, 1981
7. Горышина Т.К. Экология растений. – М.: Высшая школа,1979
Часть накопленных в процессе фотосинтеза углеводов теряется. Однако все попытки искусственно ингибировать фотодыхание приводили к общему снижению интенсивности фотосинтеза. На современном этапе развития физиологии растений принято считать, что основное значение фотодыхания заключается в его защитной роли. Сбрасывая таким образом избыточную энергию, растение избегает разрушения фотосистем, ...
У засухоустойчивых растений эти показатели будут выше. В селекционной работе используют такой показатель, как содержание статолитного крахмала в корневом чехлике. Генетически обусловленным признаком засухоустойчивости растений является способность их вегетативных органов (особенно листьев) накапливать во время засухи пролин. При этом концентрация пролина увеличивается в 10-100 раз. В пролине...
Вода, поступившая в клетки корня под влиянием разности водных потенциалов, которые возникают благодаря транспирации и корневого давления, передвигается до проводящих элементов ксилемы. Согласно современным представлениям, вода в корневой системе может перемещаться в радиальном направлении тремя путями:
- апопластическим,
- симпластическим,
- трансмембранным.
При транспорте по апопласту вода передвигается по клеточным стенкам, не проходя через мембраны. При симпластном т ранспорте вода проникает в клетку через полупроницаемую мембрану и далее перемещается по протопластам клеток, которые соединены между собой многочисленными плазмодесмами. При трансмембранном транспорте вода перетекает через клетки и при этом проходит, по крайней мере, две плазматические мембраны. В последнее время много внимания уделяется аквапоринам - мембранным белкам, образующим в мембранах специализированные водные каналы и определяющим проницаемость для воды. Эксперименты показали, что передвижение воды по коре корня идет главным образом по апопласту, где она встречает меньшее сопротивление, и лишь частично по симпласту (С. Френч). Апопластный путь прерывается в эндодерме в связи с наличием поясков Каспари . Вместе с тем в апикальной части суберинизация отсутствует, поэтому вода легко проникает через эндодерму. Кроме того, в суберинизированных частях корня вода может проходить через пропускные клетки.
Для транспорта в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану клеток эндодермы. Таким образом, мы имеем дело как бы с осмометром, у которого полупроницаемая мембрана расположена в клетках эндодермы. Вода устремляется через эту мембрану. В сторону меньшего (более отрицательного) водного потенциала. Далее вода поступает в сосуды ксилемы.
По вопросу о причинах, вызывающих секрецию воды в сосуды ксилемы, имеются различные суждения. Согласно гипотезе Крафтса , это следствие выброса солей в сосуды ксилемы, в результате чего там создается повышенная их концентрация, и водный потенциал становится более отрицательным . Предполагается, что в результате активного поступления соли накапливаются в клетках корня. Однако интенсивность дыхания в клетках, окружающих сосуды ксилемы (перицикл), очень низкая, и они не удерживают соли, которые благодаря этому десорбируются в сосуды. Транспорт воды в корне зависит от интенсивности процесса дыхания . При помещении растений в условия, тормозящие дыхание корней (низкая температура, анаэробиоз или наличие дыхательных ядов), они транспортируют меньше воды.
Дальнейшее передвижение воды идет по сосудистой системе корня, стебля и листа. Проводящие элементы ксилемы состоят из сосудов и трахеид. Опыты с кольцеванием показали, что восходящий ток воды по растению движется в основном по ксилеме. В проводящих элементах ксилемы вода встречает незначительное сопротивление, что, естественно, облегчает передвижение воды на большие расстояния. Правда, некоторое количество воды передвигается и вне сосудистой системы. Вне ксилемы движется всего от 1 до 10% общего потока воды. Из сосудов стебля вода попадает в сосуды листа. Вода движется из стебля через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся все более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Именно поэтому густота жилкования листа считается одним из важнейших признаков ксероморфной структуры - отличительной чертой растений, устойчивых к засухе.
Вся вода в клетке находится в равновесном состоянии . Иначе говоря, в смысле насыщенности водой имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. В связи с этим, как только в силу процесса транспирации возникает ненасыщенность водой клеточных стенок паренхимных клеток, она сейчас же передается внутрь клетки, водный потенциал которой падает. Вода передвигается от клетки к клетке благодаря градиенту водного потенциала . По-видимому, передвижение воды от клетки к клетке в листовой паренхиме идет не по симпласту, а в основном по клеточным стенкам , где сопротивление значительно меньше. По сосудам вода движется благодаря создающемуся в силу транспирации градиенту водного потенциала, градиенту свободной энергии (от системы с большей свободой энергии к системе с меньшей). Распределение водных потенциалов, которое и вызывает передвижение воды: водный потенциал почвы (-0,5 бар), корня (-2 бар), стебля (-5 бар), листьев (-15 бар), воздуха при относительной влажности 50% (-1000 бар).
Восходящий ток воды может осуществляться и при отсутствии транспирации. К этому же выводу приводят опыты, показывающие ритмическое секретирование устьичными клетками жидкой воды, а также зависимость передвижения воды от эндогенной энергии, поставляемых процессом дыхания. Это позволяет считать, что движущая сила транспорта воды в растении является суммой двух весьма различных по своей природе составляющих, условно названных метаболической и осмотической . Осмотическая составляющая представлена в корнях сугубо осмотическими явлениями, в стебле и листьях - гидростатической тягой, создаваемой градиентом водного потенциала в системе почва - растение - атмосфера. Температурный коэффициент Q этой составляющей близок к единице, что характерно для простых физических процессов . В то же время Q процесса транспорта воды значительно выше. Столь высокий температурный коэффициент свойственен сложным процессам цепного характера. Очевидно, транспорт воды за счет метаболической составляющей относится к разряду именно таких процессов. Согласно имеющимся экспериментальным данным, в формировании метаболической составляющей непосредственное участие принимают контрактильные системы паренхимных клеток (т. е. белки цитоскелета). Они могут играть роль сфинктеров, регулирующих просветы водных каналов (открывание-закрывание) в плазмодесмах. Благодаря их работе индуцируются ритмические микроколебания внутриклеточного (внутрисимпластного) гидростатического давления, которые внешне выражаются в короткопериодных (1-3 мин) автоколебаниях поступления воды в клетку (симпласт, ткань, орган) и водоотдачи (т. е. микропульсациях). Итак, процесс транспорта воды состоит из двух последовательных, ритмично чередующихся, относительно самостоятельных, хотя и тесно взаимосвязанных фаз:
- фазы сокращения, во время которой вода выделяется (выталкивается, секретируется),
- последующей фазы расслабления, во время которой вода поглощается.
Выделение воды вызывает падение тургора и водного потенциала в целом, создавая предпосылку для поглощения следующей порции воды, вновь приводящего к возрастанию водного потенциала вплоть до того, что он из отрицательного становится положительным. После этого происходит новое сокращение. Именно фаза сокращения происходит с участием контрактильных систем и требует затраты энергии. Таким образом, вода поглощается и выделяется по градиенту водного потенциала , транспорт воды в термодинамическом понимании является пассивным .
Возникающие за счет ритмической деятельности внутриклеточного сократительного аппарата микроколебания гидростатического давления паренхимных клеток являются механизмом, создающим локальные градиенты водного потенциала на пути водного тока и тем самым регулирующим скорость этого тока. Именно благодаря этому формируется метаболическая составляющая движущей силы транспорта воды в растении, играющая решающую роль в общей системе эндогенной регуляции. Под влиянием ингибиторов контрактильных систем или окислительного фосфорилирования (т. е. при нарушении энергоснабжения) противофазность исчезает, автоколебания затухают и транспорт воды тормозится (В.Н. Жолкевич).
Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту, значительно большую 10 м. Расчеты показали, что благодаря наличию сродства между молекулами воды силы сцепления достигают величины -30 бар. Это такая сила, которая позволяет поднять воду на высоту, равную 120 м, без разрыва водных нитей, что примерно составляет максимальную высоту деревьев. Силы сцепления существуют и между водой и стенками сосудов - адгезия . Стенки проводящих элементов ксилемы эластичны. Даже при недостатке воды связь между молекулами воды и стенками сосудов не нарушается. Это подтверждается исследованиями по изменению толщины стебля травянистых растений. Определения показали, что в полуденные часы толщина стебля растений уменьшается. Если перерезать стебель, то сосуды сразу расширяются и воздух поступает в них.
Степень натяжения водных нитей в сосудах зависит от соотношения процессов поглощения и испарения воды. Все это позволяет растительному организму поддерживать единую водную систему и не обязательно восполнять каждую каплю испаряемой воды. Таким образом, при нормальном водоснабжении создается непрерывность воды в системе почва - растение - атмосфера. В том случае, если в отдельные членики сосудов попадает воздух (эмболия), они, по-видимому, выключаются из общего тока проведения воды. Таков путь воды по растению и его основные движущие силы.
Филогенетически корень возник позже стебля и листа - в связи с переходом растений к жизни на суше и вероятно, произошёл от корнеподобных подземных веточек. У корня нет ни листьев, ни в определённом порядке расположенных почек. Для него характерен верхушечный рост в длину, боковые разветвления его возникают из внутренних тканей, точка роста покрыта корневым чехликом. Корневая система формируется на протяжении всей жизни растительного организма. Иногда корень может служить местом отложения в запас питательных веществ. В таком случае он видоизменяется.
Виды корней
Главный корень образуется из зародышевого корешка при прорастании семени. От него отходят боковые корни.
Придаточные корни развиваются на стеблях и листьях.
Боковые корни представляют собой ответвления любых корней.
Каждый корень (главный, боковые, придаточные) обладает способностью к ветвлению, что значительно увеличивает поверхность корневой системы, а это способствует лучшему укреплению растения в почве и улучшению его питания.
Типы корневых систем
Различают два основных типа корневых систем: стержневая, имеющая хорошо развитый главный корень, и мочковатая. Мочковатая корневая система состоит из большого числа придаточных корней, одинаковых по величине. Вся масса корней состоит из боковых или придаточных корешков и имеет вид мочки.
Сильно разветвлённая корневая система образует огромную поглощающую поверхность. Например,
- общая длина корней озимой ржи достигает 600 км;
- длина корневых волосков — 10 000 км;
- общая поверхность корней — 200 м 2 .
Это во много раз превышает площадь надземной массы.
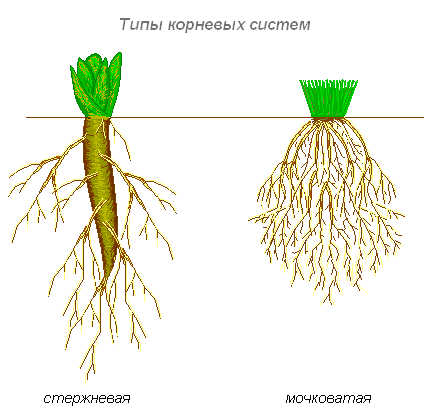
Если у растения хорошо выражен главный корень и развиваются придаточные корни, то формируется корневая система смешанного типа (капуста, помидор).
Внешнее строение корня. Внутреннее строение корня
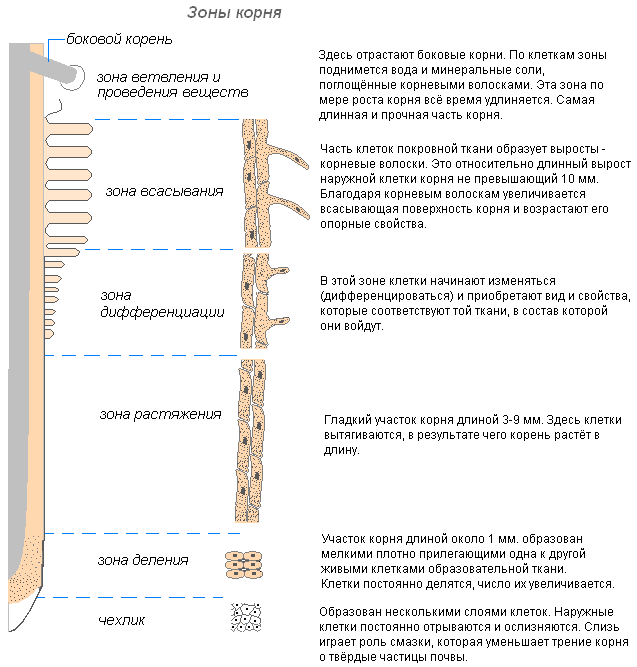
Зоны корня
Корневой чехлик
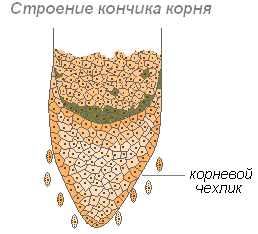
Корень растёт в длину своей верхушкой, где находятся молодые клетки образовательной ткани. Растущая часть покрыта корневым чехликом, защищающим кончик корня от повреждений, и облегчает продвижение корня в почве во время роста. Последняя функция осуществляется благодаря свойству внешних стенок корневого чехлика покрываться слизью, что уменьшает трение между корнем и частичками почвы. Могут даже раздвигать частички почвы. Клетки корневого чехлика живые, часто содержат зёрна крахмала. Клетки чехлика постоянно обновляются за счёт деления. Участвует в положительных геотропических реакциях (направление роста корня к центру Земли).
Клетки зоны деления активно делятся, протяженность этой зоны у разных видов и у разных корней одного и того же растения неодинакова.
За зоной деления расположена зона растяжения (зона роста). Протяжённость этой зоны не превышает нескольких миллиметров.
По мере завершения линейного роста наступает третий этап формирования корня — его дифференциация, образуется зона дифференциации и специализации клеток (или зона корневых волосков и всасывания). В этой зоне уже различают наружный слой эпиблемы (ризодермы) с корневыми волосками, слой первичной коры и центральный цилиндр.
Строение корневого волоска
Корневые волоски — это сильно удлинённые выросты наружных клеток, покрывающих корень. Количество корневых волосков очень велико (на 1 мм 2 от 200 до 300 волосков). Их длина достигает 10 мм. Формируются волоски очень быстро (у молодых сеянцев яблони за 30-40 часов). Корневые волоски недолговечны. Они отмирают через 10-20 дней, а на молодой части корня отрастают новые. Это обеспечивает освоение корнем новых почвенных горизонтов. Корень непрерывно растёт, образуя всё новые и новые участки корневых волосков. Волоски могут не только поглощать готовые растворы веществ, но и способствовать растворению некоторых веществ почвы, а затем всасывать их. Участок корня, где корневые волоски отмерли, некоторое время способен всасывать воду, но затем покрывается пробкой и теряет эту способность.

Оболочка волоска очень тонкая, что облегчает поглощение питательных веществ. Почти всю клетку волоска занимает вакуоль, окружённая тонким слоем цитоплазмы. Ядро находится в верхней части клетки. Вокруг клетки образуется слизистый чехол, который содействует склеиванию корневых волосков с частицами почвы, что улучшает их контакт и повышает гидрофильность системы. Поглощению способствует выделение корневыми волосками кислот (угольной, яблочной, лимонной), которые растворяют минеральные соли.
Корневые волоски играют и механическую роль — они служат опорой верхушке корня, которая проходит между частичками почвы.
Под микроскопом на поперечном срезе корня в зоне всасывания видно его строение на клеточном и тканевом уровнях. На поверхности корня — ризодерма, под ней — кора. Наружный слой коры — экзодерма, вовнутрь от неё — основная паренхима. Её тонкостенные живые клетки выполняют запасающую функцию, проводят растворы питательных веществ в радиальном направлении — от всасывающей ткани к сосудам древесины. В них же происходит синтез ряда жизненно важных для растения органических веществ. Внутренний слой коры — эндодерма. Растворы питательных веществ, поступающие из коры в центральный цилиндр через клетки эндодермы, проходят только через протопласт клеток.

Кора окружает центральный цилиндр корня. Она граничит со слоем клеток, долго сохраняющих способность к делению. Это перицикл. Клетки перицикла дают начало боковым корням, придаточным почкам и вторичным образовательным тканям. Вовнутрь от перицикла, в центре корня, находятся проводящие ткани: луб и древесина. Вместе они образуют радиальный проводящий пучок.
Проводящая система корня проводит воду и минеральные вещества из корня в стебель (восходящий ток) и органические вещества из стебля в корень (нисходящий ток). Состоит она из сосудисто-волокнистых пучков. Основными слагаемыми частями пучка являются участки флоэмы (по ним вещества передвигаются к корню) и ксилемы (по которым вещества передвигаются от корня). Основные проводящие элементы флоэмы — ситовидные трубки, ксилемы — трахеи (сосуды) и трахеиды.
Процессы жизнедеятельности корня
Транспорт воды в корне
Всасывание воды корневыми волосками из почвенного питательного раствора и проведение её в радиальном направлении по клеткам первичной коры через пропускные клетки в эндодерме к ксилеме радиального проводящего пучка. Интенсивность поглощения воды корневыми волосками называется сосущей силой (S), она равна разнице между осмотическим (P) и тургорным (T) давлением: S=P-T.
Когда осмотическое давление равно тургорному (P=T), то S=0, вода перестаёт поступать в клетку корневого волоска. Если концентрация веществ почвенного питательного раствора будет выше, чем внутри клетки, то вода будет выходить из клеток и наступит плазмолиз — растения завянут. Такое явление наблюдается в условиях сухости почвы, а также при неумеренном внесении минеральных удобрений. Внутри клеток корня сосущая сила корня возрастает от ризодермы по направлению к центральному цилиндру, поэтому вода движется по градиенту концентрации (т. е. из места с большей её концентрацией в место с меньшей концентрацией) и создаёт корневое давление, которое поднимает столбик воды по сосудам ксилемы, образуя восходящий ток. Это можно обнаружить на весенних безлистных стволах, когда собирают «сок», или на срезанных пнях. Истекание воды из древесины, свежих пней, листьев, называется «плачем» растений. Когда распускаются листья, то они тоже создают сосущую силу и притягивают воду к себе — образуется непрерывный столбик воды в каждом сосуде — капиллярное натяжение. Корневое давление является нижним двигателем водного тока, а сосущая сила листьев — верхним. Подтвердить это можно с помощью несложных опытов.
Всасывание воды корнями
Цель: выяснить основную функцию корня.
Что делаем: растение, выращенное на влажных опилках, отряхнём его корневую систему и опустим в стакан с водой его корни. Поверх воды для защиты её от испарения нальём тонкий слой растительного масла и отметим уровень.

Что наблюдаем: через день-два вода в ёмкости опустилась ниже отметки.

Результат: следовательно, корни всосали воду и подали её наверх к листьям.
Можно ещё проделать один опыт, доказывающий всасывание питательных веществ корнем.
Что делаем: срежем у растения стебель оставив пенёк высотой 2-3 см. На пенёк наденем резиновую трубку длиной 3 см, а на верхний конец наденем изогнутую стеклянную трубку высотой 20-25 см.
Что наблюдаем: вода в стеклянной трубке поднимается, и вытекает наружу.
Результат: это доказывает, что воду из почвы корень всасывает в стебель.

А влияет ли температура воды на интенсивность всасывания корнем воды?
Цель: выяснить, как температура влияет на работу корня.
Что делаем: один стакан должен быть с тёплой водой (+17-18ºС), а другой с холодной (+1-2ºС).
Что наблюдаем: в первом случае вода выделяется обильно, во втором — мало, или совсем приостанавливается.
Результат: это является доказательством того, что температура сильно влияет на работу корня.

Тёплая вода активно поглощается корнями. Корневое давление повышается.

Холодная вода плохо поглощается корнями. В этом случае корневое давление падает.
Минеральное питание
Физиологическая роль минеральных веществ очень велика. Они являются основой для синтеза органических соединений, а также факторами, которые изменяют физическое состояние коллоидов, т.е. непосредственно влияют на обмен веществ и строение протопласта; выполняют функцию катализаторов биохимических реакций; воздействуют на тургор клетки и проницаемость протоплазмы; являются центрами электрических и радиоактивных явлений в растительных организмах.
Установлено, что нормальное развитие растений возможно только при наличии в питательном растворе трёх неметаллов — азота, фосфора и серы и — и четырёх металлов — калия, магния, кальция и железа. Каждый из этих элементов имеет индивидуальное значение и не может быть заменён другим. Это макроэлементы, их концентрация в растении составляет 10 -2 –10%. Для нормального развития растений нужны микроэлементы, концентрация которых в клетке составляет 10 -5 –10 -3 %. Это бор, кобальт, медь, цинк, марганец, молибден др. Все эти элементы есть в почве, но иногда в недостаточном количестве. Поэтому в почву вносят минеральные и органические удобрения.
Растение нормально растёт и развивается в том случае, если в окружающей корни среде будут содержаться все необходимые питательные вещества. Такой средой для большинства растений является почва.
Дыхание корней
Для нормального роста и развития растения необходимо чтобы к корню поступал свежий воздух. Проверим, так ли это?
Цель: нужен ли воздух корню?
Что делаем: возьмём два одинаковых сосуда с водой. В каждый сосуд поместим развивающие проростки. Воду в одном из сосудов каждый день насыщаем воздухом с помощью пульверизатора. На поверхность воды во втором сосуде нальём тонкий слой растительного масла, так как оно задерживает поступление воздуха в воду.

Что наблюдаем: через некоторое время растение во втором сосуде перестанет расти, зачахнет, и в конце концов погибнет.

Результат: гибель растения наступает из-за недостатка воздуха, необходимого для дыхания корня.
Видоизменения корней
У некоторых растений в корнях откладываются запасные питательные вещества. В них накапливаются углеводы, минеральные соли, витамины и другие вещества. Такие корни сильно разрастаются в толщину и приобретают необычный внешний вид. В формировании корнеплодов участвуют и корень, и стебель.
Корнеплоды
Если запасные вещества накапливаются в главном корне и в основании стебля главного побега, образуются корнеплоды (морковь). Растения, образующие корнеплоды, в основном двулетники. В первый год жизни они не цветут и накапливают в корнеплодах много питательных веществ. На второй — они быстро зацветают, используя накопленные питательные вещества и образуют плоды и семена.
Корневые клубни

У георгина запасные вещества накапливаются в придаточных корнях, образуя корневые клубни.
Бактериальные клубеньки
Своеобразно изменены боковые корни у клевера, люпина, люцерны. В молодых боковых корешках поселяются бактерии, что способствует усвоению газообразного азота почвенного воздуха. Такие корни приобретают вид клубеньков. Благодаря этим бактериям эти растения способны жить на бедных азотом почвах и делать их более плодородными.
Ходульные

У пандуса, произрастающего в приливно-отливной зоне, развиваются ходульные корни. Они высоко над водой удерживают на зыбком илистом грунте крупные облиственные побеги.
Воздушные

У тропических растений, живущих на ветвях деревьев, развиваются воздушные корни. Они часто встречаются у орхидей, бромелиевых, у некоторых папоротников. Воздушные корни свободно висят в воздухе, не достигая земли и поглощая попадающую на них влагу от дождя или росы.
Втягивающие

У луковичных и клубнелуковичных растений, например у крокусов, среди многочисленных нитевидных корней имеется несколько более толстых, так называемых втягивающих, корней. Сокращаясь, такие корни втягивают клубнелуковицу глубже в почву.
Столбовидные

У фикуса развиваются столбовидные надземные корни, или корни-подпорки.
Почва как среда обитания корней
Почва для растений является средой, из которой оно получает воду и элементы питания. Количество минеральных веществ в почве зависит от специфических особенностей материнской горной породы, деятельности организмов, от жизнедеятельности самих растений, от типа почвы.
Почвенные частицы конкурируют с корнями за влагу, удерживая её своей поверхностью. Это так называемая связанная вода, которая подразделяется на гигроскопическую и плёночную. Удерживается она силами молекулярного притяжения. Доступная растению влага представлена капиллярной водой, которая сосредоточена в мелких порах почвы.
Между влагой и воздушной фазой почвы складываются антагонистические отношения. Чем больше в почве крупных пор, тем лучше газовый режим этих почв, тем меньше влаги удерживает почва. Наиболее благоприятный водно-воздушный режим поддерживается в структурных почвах, где вода и воздух находятся одновременно и не мешают друг другу — вода заполняет капилляры внутри структурных агрегатов, а воздух — крупные поры между ними.
Характер взаимодействия растения и почвы в значительной степени связан с поглотительной способностью почвы — способностью удерживать или связывать химические соединения.
Микрофлора почвы разлагает органические вещества до более простых соединений, участвует в формировании структуры почвы. Характер этих процессов зависит от типа почвы, химического состава растительных остатков, физиологических свойств микроорганизмов и других факторов. В формировании структуры почвы принимают участие почвенные животные: кольчатые черви, личинки насекомых и др.
В результате совокупности биологических и химических процессов в почве образуется сложный комплекс органических веществ, который объединяют термином «гумус».
Метод водных культур
В каких солях нуждается растение, и какое влияние оказывают они на рост и развитие его, было установлено на опыте с водными культурами. Метод водных культур — это выращивание растений не в почве, а в водном растворе минеральных солей. В зависимости от поставленной цели в опыте можно исключить отдельную соль из раствора, уменьшить или увеличить ее содержание. Было выяснено, что удобрения, содержащие азот, способствуют росту растений, содержащие фосфор — скорейшему созреванию плодов, а содержащие калий — быстрейшему оттоку органических веществ от листьев к корням. В связи с этим содержащие азот удобрения рекомендуется вносить перед посевом или в первой половине лета, содержащие фосфор и калий — во второй половине лета.
С помощью метода водных культур удалось установить не только потребность растения в макроэлементах, но и выяснить роль различных микроэлементов.
В настоящее время известны случаи, когда выращивают растения методами гидропоники и аэропоники.
Гидропоника — выращивание растений в сосудах, заполненных гравием. Питательный раствор, содержащий необходимые элементы, подаётся в сосуды снизу.
Аэропоника — это воздушная культура растений. При этом способе корневая система находится в воздухе и автоматически (несколько раз в течение часа) опрыскивается слабым раствором питательных солей.









